3 Results and Discussion
There are 331 articles for the term “landscape metrics”, 131 articles for the term “landscape indices”, and 17 for “landscape indexes” in the ISI Web of Science literature database (October 14, 2008). In fact, some of the articles overlapped, and a few articles were added by using the singular form of the two terms. We therefore eventually reviewed 337 articles for the term “landscape metrics”, and 141 articles for the term “landscape indices” and “landscape indexes”.
3.1 “Landscape metrics” vs “landscape indices”
The term “landscape metrics” appears more frequently, but there are no definite rules or traditions as to when one or the other term is used. It seems that the term “landscape metrics” is used more often only for metrics calculated by Fragstats or by some of its developments in other programs (FRAG*ARC, Fragstats for ArcView, Patch Analyst etc). The term “landscape indices” is more frequently used in a broader sense, i.e., metrics have similar or the same formulae as in Fragstats but are named differently. Also, in terms of the timeline of their use (Figure 1*), it can be seen that the term “landscape indices” was more often used in the 1990s, and now the term “landscape metrics” is prevalent. From here on we will only use the term “landscape metrics”.
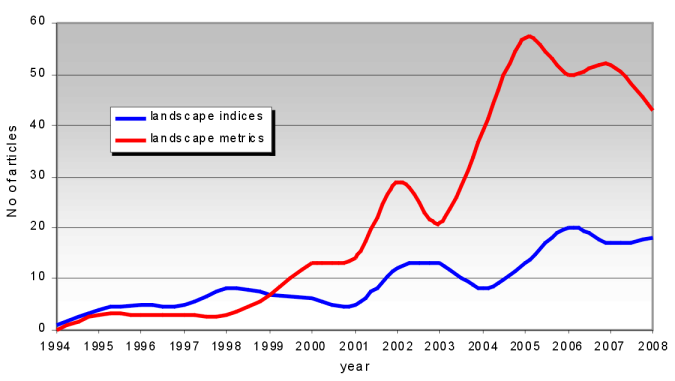
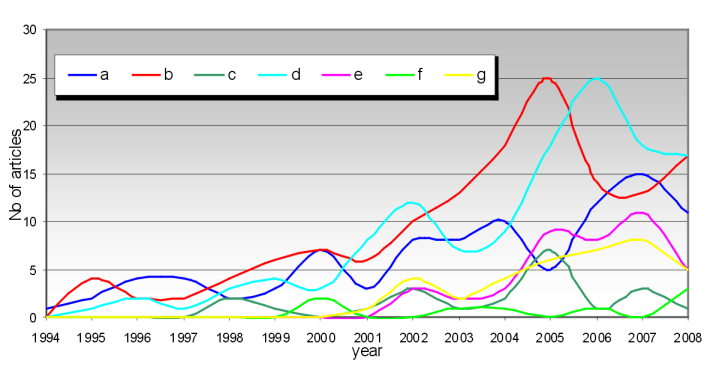
The usage of landscape metrics is very broad, and can be grouped into seven general categories: a) use and misuse/ selection of metrics; b) biodiversity and habitat analysis; c) estimating water quality; d) evaluation of landscape pattern and changes therein; e) urban landscape pattern, road network; f) aesthetics of landscape; g) management, planning and monitoring. Category e) is actually more like a subcategory of d), but since the human impact on landscapes is so important and has been very widely explored, we decided to analyze it separately. Most of the studies published from 1994 – 2008 are on biodiversity and habitat analysis, and the evaluation of landscape pattern and changes therein (up to 25 articles per year; Figure 2*). There are up to 15 articles per year about the use and misuse / selection of metrics. The number of articles published each year has generally increased since 1994, but for example articles published on the relations between landscape aesthetics relations and landscape metrics has been surprisingly low.
3.2 Use and selection of metrics
Many of the articles have been written on the use and misuse and selection principles of landscape metrics. As there are literally hundreds of landscape metrics, many of the landscape metrics are correlated to each other, which makes interpretation more difficult. Riitters et al. (1995) found that the first six factors explained about 87% of the variation in the 26 landscape metrics, and these factors were interpreted as composite measures of average patch compaction, overall image texture, average patch shape, patch perimeter-area scaling, the number of attribute classes, and large-patch density-area scaling. Cushman et al. (2008) performed principal component analysis (PCA) and cluster analysis to identify independent components of landscape structure and group them, and found that there were eight universal and consistent combinations of Fragstats metrics that universally describe the major attributes of landscape structure at the landscape level. Botequilha Leitão and Ahern (2002*) even proposed a core set of metrics that are most useful and relevant for landscape planning and Schindler et al. (2008) proposed set of metrics for establishing a landscape monitoring program, to detect the local drivers of biodiversity in Mediterranean area. The interpretation of some of the landscape metrics is also complicated because their behaviour has not yet been evaluated (McGarigal et al., 2002). In that field, neutral landscapes are useful (see for example Neel et al., 2004; Li et al., 2005a).
Another important issue is that the results of spatial data analysis depend on data aggregation methods and the zoning scheme. Its general formulation is known as the modifiable areal unit problem (MAUP), created by (Openshaw and Taylor, 1981). MAUP in the context of landscape ecology consists of three related aspects: how the grain size, zoning and areal extent of investigation influence results, and how to determine their optimal values for each particular case. The dependence of landscape metrics on grain size is studied by (Wickham and Riitters, 1995; Uuemaa et al., 2005*; Buyantuyev and Wu, 2007). Wickham et al. (1997); Huang et al. (2006); Langford et al. (2006) have studied the influence of map classification, and Wu et al. (2002) and Wu (2004) have studied the extent of the study area on the value of landscape metrics.
Although there are already hundreds of landscape metrics, several researchers have proposed new landscape metrics. Jaeger (2000) proposed a degree of landscape division (D), splitting index (S), and effective mesh size (m), which characterize the anthropogenic penetration of landscapes from a geometric point of view and are calculated from the distribution function of the remaining patch sizes. (He et al., 2000) proposed new metric (aggregation index-AI) for measuring aggregation in landscape pattern. AI is class-based, and contrary to contagion, it is independent of composition. All of these metrics are now also available in Fragstats. The fact that researchers are still working out new metrics indicates the need for measuring new aspects of landscape pattern but also researchers’ aspiration to overcome the collinearity in Fragstats metrics.
3.3 Biodiversity and habitat analysis
The relationship between landscape metrics and bird species richness and their habitat preferences has been studied most extensively. There are many studies that have been performed on the relationships between landscape parameters and specific species – owls (Carey et al., 1992; Ribe et al., 1998), sparrows (Perkins and Conner, 2003), turkeys (Chamberlain et al., 2000; Miller and Conner, 2007), woodpeckers (Wigley et al., 1999), bobwhites (Guthery et al., 2001; Twedt et al., 2007), grouses (Fearer and Stauffer, 2003), pheasants (Clark et al., 1999) and ducks (Stephens et al., 2005). Different studies have shown that most bird species responded more strongly to the composition of land-cover classes than to the configuration of the landscape (Table 1). Of landscape configuration metrics, patch size has given the most important relationships with bird species richness, i.e., fragmentation plays an important role for birds.
Landscape metrics have been used to determine the landscape preferences of raccoons (Henner et al., 2004), gray wolves (Mladenoff et al., 1995), wild hogs (Gaines et al., 2005*); moose (Maier et al., 2005*), deer (Foster et al., 1997*; Finder et al., 1999; Kie et al., 2002), black bears (Kindall and Van Manen, 2007), ocelots (Jackson et al., 2005*), elk (Stubblefield et al., 2006), possums (Eyre and Buck, 2005*) and bats (Limpert et al., 2007). Different species provide different correlations with landscape metrics depending on their landscape preferences, i.e., large compact patches are preferred by wild hogs (Gaines et al., 2005), moose (Maier et al., 2005), deer (Table 1; Foster et al., 1997*; Plante et al., 2004*) and possums (Eyre and Buck, 2005), while ocelots (Jackson et al., 2005) and gliders (Table 1; McAlpine and Eyre, 2002*) preferred areas that had a greater degree of fragmentation (i.e., a larger number of patches of smaller size, and with more edge).
Several studies have even shown the significance of landscape pattern (measured by landscape metrics) on frog populations (Table 1; Knutson et al., 1999; Pellet et al., 2004*) and on insect population levels (Radeloff et al., 2000*; French et al., 2004; Roschewitz et al., 2005). The structure of the landscape may be important to explain and understand the epidemiology associated with insects (Graham et al., 2004). Overgaard et al. (2003*) studied the influence of landscape structure on Anopheline mosquito density, and based on their results suggested that if landscape management were to be used for malaria control, the large-scale reduction and fragmentation of forest cover would be needed in the case of northern Thailand. The risk of transmission of Lyme disease has been found to be influenced by landscape structure and the spatial arrangement of land cover types (Turner, 1989). Brownstein et al. (2005) performed an analysis of the landscape pattern of forest patches using satellite imagery, and calculated landscape indices revealed a positive link between fragmentation and both tick density and the prevalence of infection in ticks. Yang et al. (2008) investigated ecological variability related to the distribution of Oncomelania hupensis, the snail intermediate host of Schistosoma japonicum, and found that the reduction of the heterogeneity of the landscape could reduce snail density.
It is known that habitats composed of spatially heterogeneous abiotic conditions provide a great diversity of potentially suitable niches for plant species. The scientific premises of landscape ecology suggest that, at a higher spatial level, the composition and structure of the landscape mosaic also influences biotic processes and hence species richness (Honnay et al., 2003). There are several studies on the determining of relationships between landscape structure (estimated with landscape metrics) and plant diversity (Table 1; Moser et al., 2002*; Burton and Samuelson, 2008*; Hernández-Stefanoni and Dupuy, 2008). A growing number of ecological studies have focused on alien plants’ invasive processes in order to predict further invasions and associated potentially negative effects. The introduction of alien plants or plant species new to an area due to human activity and their spread and establishment is thought to cause a decline in the diversity of native species (Williamson, 1999). Deutschewitz et al. (2003) found that the species richness of native and alien plants increases with moderate levels of natural and/or anthropogenic disturbances, coupled with high levels of habitat and structural heterogeneity in urban, riverine, and small-scale rural ecosystems. Kumar et al. (2006*) found that both native and non-native plant species richness were positively correlated with edge density, Simpson’s diversity index and the interspersion/juxtaposition index, and were negatively correlated with mean patch size (Table 1). Landscape metrics of watersheds have also proven to be useful for the identification of estuarine benthic conditions and degraded bottom communities (Hale et al., 2004).
| Taxa |
Landscape metrics |
Dependent variables |
Pearson r |
References |
| Amphibia |
Density of urban landcover in a buffer of 30 m around the pond |
calling sites selection by males of tree frog (Hyla arborea) |
−0.23*** | |
| Mammals |
log10 forest patch area |
carnivores species richness |
0.69*** | |
|
|
primates species richness |
0.65*** |
|
|
|
Mean woods patch size (log) |
deer vulnerability to harvest (log) |
−0.75**** | ||
|
Proximity index (log) |
|
−0.76**** | ||
|
mean woods patch shape (log) |
|
−0.7**** | ||
|
Mean nearest neighbour distance |
|
0.76**** | ||
|
Number of patches |
Deer density |
0.43* | ||
|
Edge density |
|
0.37*** | ||
|
Mean Patch Size |
Count of the Yellow-bellied Glider (Petaurus australis) |
−0.25* | ||
|
Edge density |
|
0.29* | ||
|
Contrast weighted edge density |
|
0.28* | ||
|
Largest patch index |
Diversity of exudivore species |
−0.26* | ||
|
Shannon’s evenness index |
|
0.33* | ||
|
Contagion |
|
−0.33* | ||
| Birds |
Agricultural land (ha) |
Number of hooded crane |
0.79** | |
|
Proportion forest |
Abundance of female cowbirds |
0.62** | ||
|
Proportion forest |
Abundance of indigo buntings |
−0.77** | ||
|
Proximity index |
|
−0.35** | ||
|
Edge density |
Abundance of wood thrushes |
0.37** | ||
| Insects |
Total area of host patches |
Mean abundance of (Delphacodes kuscheli) |
0.79*** | |
|
Mean proximity index (host patches) |
|
0.96*** | ||
|
Class area (maize) |
Total number of western corn rootworm |
0.91* | ||
|
Number of patches (maize) |
|
0.9* | ||
|
Proximity (maize patches) |
|
0.99** | ||
|
Edge density |
Mean jack pine budworm (Choristoneura pinus pinus) population levels |
0.26* | ||
|
|
Corrected mean perimeter area ratio |
0.34* | ||
|
Modified Simpson’s diversity index |
Anopheline species diversity (Shannon–Weaver diversity index) |
−0.74* | ||
| Plants |
Forest cover (%) |
Riparian woody plant species richness |
0.78** | |
|
Shannon’s diversity index |
|
−0.74** | ||
|
Number of patches |
Species richness of vascular plants |
0.61** | ||
|
Edge density |
|
0.56** | ||
|
Mean shape index |
|
−0.7** | ||
|
Number of patches |
Species richness of bryophytes |
0.6** | ||
|
Edge density |
|
0.57** | ||
|
Mean shape index |
|
−0.51** | ||
|
Simpson’s diversity index |
Native species richness |
0.28* | ||
|
Interspersion and juxtaposition index |
|
0.31* | ||
|
Patch richness density |
|
0.35* | ||
|
Simpson’s diversity index |
Non-native species richness |
0.43* | ||
|
Interspersion and juxtaposition index |
|
0.29* | ||
|
Patch richness density |
|
0.4* | ||
3.4 Estimating water quality
Landscape structure is one of the most important factors influencing nutrient and organic matter runoff in watersheds (Turner et al., 2003; Wickham et al., 2003; Uuemaa et al., 2007*). Therefore there is increasing demand for indicators and methods that make it possible to evaluate the landscape factors influencing water quality in freshwater management (Griffith, 2002). Several studies have attempted to determine the relationship between land use/land cover structure and water quality but most studies have largely relied on compositional landscape metrics (Kearns et al., 2005). It is, however, clearly important to understand not only the total area of sources and sinks in the landscape, but also their spatial arrangement relative to flowpaths (Gergel, 2005). The importance of the spatial arrangement of land cover within watersheds on water quality has been studied by Jones et al. (2001); King et al. (2005*); Li et al. (2005b*); Snyder et al. (2005); Xiao and Ji (2007*); Uuemaa et al. (2005*, 2007); see Table 2.
|
Landscape metrics |
Dependent variables |
Pearson r or Spearman ρ |
References |
|
Cropland (%) |
Log (NO3-N) |
0.67* |
|
|
Canal line density |
Soluble reactive phosphorus reduction |
0.8* |
|
|
Canal connectivity |
|
0.83* |
|
|
Canal circuity |
|
0.85* |
|
|
Canal line density |
Total nitrogen reduction |
0.84* |
|
|
Canal connectivity |
|
0.92* |
|
|
Canal circuity |
|
0.94* |
|
|
Edge density (forest) |
Conductivity |
−0.34* |
|
|
Patch density |
|
0.33* |
|
|
Contagion |
|
−0.49* |
|
|
Edge density (forest) |
Total Cd |
−0.53* |
|
|
Edge density (forest) |
Total Zn |
−0.49* |
|
|
Contagion |
|
−0.36* |
|
|
Patch density |
BOD7 (biological oxygen demand) |
−0.53* |
|
|
Edge density |
|
−0.47* |
|
|
Patch density |
CODKMnO4 (chemical oxygen demand) |
−0.49* |
|
|
Mean shape index |
|
0.66* |
|
|
Edge density |
Total-N |
−0.56* |
|
|
Mean shape index |
|
−0.44* |
|
The spatial pattern of riparian zones is also an especially powerful landscape indicator for water quality, because the variation in length, width, and gaps of riparian buffers influences their effectiveness as nutrient sinks (Gergel et al., 2002). Weller et al. (1998) developed and analyzed models predicting landscape discharge based on material release by an uphill source area, the spatial distribution of a riparian buffer along a stream, and retention within the buffer, and found average width to be the best predictor of landscape discharge for unretentive buffers. Baker et al. (2006) quantified the effects of riparian buffers on watershed nutrient discharges by using, in addition to traditional fixed-distance measures, mean buffer width, gap frequency, and measures of variation in buffer width using both “unconstrained” metrics and “flow-path” metrics constrained by surface topography.
3.5 Evaluation of landscape pattern and changes therein
Land use changes are mostly caused by humans, but also by natural disturbances. For example, Lin et al. (2006) used landscape metrics and spatial autocorrelation to assess how earthquakes and typhoons affect landscape patterns, and found that the disturbances produced variously fragmented patches, interspersed with other patches and isolated from patches of the same type across the entire Chenyulan watershed in Taiwan. Results of fire disturbances studies Keane et al. (1999) and Hudak et al. (2004) found that fire creates more diverse, fragmented and disconnected landscapes, and Kashian et al. (2004) found that large, stand-replacing fires may result in heterogeneous forest landscapes rather than homogenous forests of uniform structure. According to Viedma et al. (2006), on the contrary, fire had made the landscape less fragmented and more continuous.
Teixidó et al. (2007) used landscape metrics to study the spatial patterns of the Antarctic benthos in terms of the succession process after iceberg disturbance, and found the first stages of recovery to have low cover area, low complexity of patch shape, small patch size, low diversity and patches that were poorly interspersed to samples from later stages with higher values of these indices.
Many studies have been done on the mapping of forest cover change. Human influence also causes forest fragmentation that also affects species richness (Fuller, 2001; Cayuela et al., 2006; Echeverría et al., 2007*; Altamirano et al., 2007). Twentieth century management activities have significantly influenced the structure of the forest landscape (Wolter and White, 2002; Löfman and Kouki, 2003), and altered spatial patterns of physiognomies, cover types and structural conditions, and vulnerabilities to fire, insect, and pathogen disturbances (Hessburg et al., 2000). Logging is one of the main reasons for forest fragmentation, and although it may change the landscape structure at a small spatial scale and not alter the structure of the entire forest mosaic (Leimgruber et al., 2002), it can be associated with dramatic changes in the structure and composition of the forests (Echeverría et al., 2007). Etheridge et al. (2006) also found, using landscape metrics as an indicator, that clearcuts result in the loss of large patches. Zhang and Guindon (2005) used landscape metrics and cellular automata to analyze human impacts on forest fragmentation, and showed that the observed values of scalar landscape metrics and their interrelationships can only be understood by taking into account the spatial pattern aspects associated with causal human drivers of the deforestation process.
3.6 Urban landscape pattern, road network
Urbanization has significantly changed natural landscapes everywhere. Urban growth and fragmentation caused by urban sprawl have been extensively studied (Herold et al., 2002; Ji et al., 2006; Tang et al., 2006; Gonzalez-Abraham et al., 2007). It has been shown that landscape pattern is more fragmented around city centres and along coastlines, where urbanization and human economic activities are more concentrated (Yang and Liu, 2005). There are several possibilities as to how to use landscape metrics to detect spatial patterns caused by urbanization. For example, Seto and Fragkias (2005) calculated and analyzed landscape metrics spatiotemporally across three buffer zones, but another effective approach for analyzing systematically the effects of urbanization on ecosystems is to studying the changes in ecosystem patterns and processes along an urban-to-rural gradient (McDonnell et al., 1997). Studies of landscape pattern change along an urban-to-rural gradient focus on the identification of urban texture – whether urban landscapes have unique “spatial signatures” that are distinguishable from other types of landscapes (Weng, 2007*). In many studies only land use changes in space are considered (Luck and Wu, 2002; Hahs and McDonnell, 2006; Conway and Hackworth, 2007), but landscape pattern also changes over time. Spatiotemporal gradient analysis makes it possible to determine how the urban centre has shifted in space and time (Wu et al., 2006; Xie et al., 2006; Weng, 2007).
Another critical issue is fragmentation caused by infrastructure, and many studies have revealed that road corridors in the urban landscape increased habitat fragmentation (Saunders et al., 2002; Zhu et al., 2006; Hawbaker et al., 2006; Jaeger et al., 2007).
3.7 Landscape aesthetics
Human perception and intuition can strongly influence how we measure and interpret landscape pattern (D’Eon and Glenn, 2000). Antrop and Van Eetvelde (2000) also emphasise the importance of holism, and found that summed entropy corresponds most closely to the landscape units defined by visual image interpretation, and can be used as a quantitative characteristic of holistically defined landscape units. (Franco et al., 2003*) also found strong explanatory relationship between citizens’ scenic beauty estimation and the Shannon’s diversity index (Table 3).
|
Landscape metrics |
Dependent variables |
Pearson r or Spearman ρ |
References |
|
Shannon’s diversity index |
Scenic beauty estimation |
0.82**** |
|
|
Edge density |
Scenic beauty estimation |
0.42*** |
|
|
Agricultural and open land |
|
0.38*** |
|
|
Recreation |
|
0.31** |
|
|
Other urban |
|
−0.45*** |
|
|
Waste |
|
−0.33** |
|
|
Wetland, open water |
|
0.33** |
|
|
Largest patch index |
Neighbourhood satisfaction (0= not satisfied; 6= very satisfied) |
0.18** |
|
|
Patch density |
|
−0.13* |
|
|
Total core area |
|
0.21** |
|
|
Mean Euclidean nearest neighbour distance |
|
−0.23** |
|
|
Area-weighted mean shape index |
|
0.15** |
|
|
Mean shape index |
|
−0.21** |
|
|
Cohesion |
|
0.14* |
|
|
Shannon’s diversity index |
Preference scores (1= least preferred; 5= most preferred) |
0.58** |
|
|
No. of land types |
|
0.53** |
|
|
No. of patches |
|
0.45* |
|
|
Percent open area |
|
−0.45* |
|
Palmer (2004) investigated residents’ perceptions of scenic quality in the Cape Cod community of Dennis, Massachusetts over a period of significant landscape change, and used landscape metrics to predict residents’ perception of scenic value for each time period. The results indicated that landscape composition metrics were more closely related to scenic value than the configuration metrics (Table 3). Scenic value was most positively related to the relative amount of agricultural and related open lands, and the most intensive urban land uses were negatively associated with scenic value. From configuration metrics only edge density was positively related to scenic value. Lee et al. (2008), however, found that residents’ neighbourhood satisfaction was more likely to be high when tree patches in neighbourhood environments were less fragmented, less isolated, and well connected (Table 3). Dramstad et al. (2006) also found that spatial configuration is related to people’s landscape preferences, and these may therefore be suitable as indicators for the visual landscape (Table 3).
The embedding of the concept of valuable landscapes in legislation such as the European Landscape
Convention (Council of Europe, 2004) has led to the need for an ‘objective’ assessment of these values and
the potential impact of changes to them (Sang et al., 2008*). The visual characteristics of a landscape are
some of the most widely experienced but also most difficult, and controversial (Walker, 1995), to define.
Sang et al. (2008) investigated how well landscape metrics predict the results on preference from the
Visulands Pan European Survey ( http://www.esac.pt/visulands) and the implications of this for the role
of preference metrics and visualisation methods in planning processes such as landscape character
assessment. Furthermore, Fry et al. (2009) find that landscape metrics have a strong conceptual base in
landscape ecological principles but for the visual aspects of landscapes this conceptual base is often
missing and thus hindering progress in the development of indicators. Therefore they proposed
hierarchical framework for establishing and strengthening links between theory and indicator
application.
http://www.esac.pt/visulands) and the implications of this for the role
of preference metrics and visualisation methods in planning processes such as landscape character
assessment. Furthermore, Fry et al. (2009) find that landscape metrics have a strong conceptual base in
landscape ecological principles but for the visual aspects of landscapes this conceptual base is often
missing and thus hindering progress in the development of indicators. Therefore they proposed
hierarchical framework for establishing and strengthening links between theory and indicator
application.
3.8 Management and planning
Landscape metrics are useful for the application of the concepts of landscape ecology to sustainable landscape planning (Botequilha Leitão and Ahern, 2002) and landscape monitoring (Herzog and Lausch, 1999). Lin et al. (2007) combined a land use change model, landscape metrics and a watershed hydrological model with an analysis of the impacts of future land use scenarios on land use pattern and hydrology for a landscape management plan. Landscape metrics also make it possible to detect potential areas for greenways (Colantonio Venturelli and Galli, 2006; Zhang and Wang, 2006), and assess habitat suitability (Holzkämper et al., 2006; Kim and Pauleit, 2007) for landscape planning and management.
Landscape metrics can be useful for assessing soil erosion on large territories (Li, 2008) and help landscape managers to indicate how well landscapes function to retain, not “leak”, vital system resources such as rainwater and soil (Ludwig et al., 2002).
Agricultural activities have major effect on floral and faunal species richness of anthropogenic landscapes. As European Union is providing subsidies to farmers for environmentally friendly agricultural practices, there is an urgent need to assess the effectiveness of these subsides. Wrbka et al. (2008*) investigated the agri-environmental measures in a parcel-wise manner and analyzed their effects on landscape values and biodiversity. They found that reduction of agrochemicals showed positive effects on biodiversity of vascular plants in grassland and birds in arable land and targeted measures that directly address threatened species were most effective, but had much less coverage. Wrbka et al. (2008) also concluded that agri-environmental measures are currently not targeted enough to effectively halt biodiversity losses. Therefore we also find that researchers should aim for more specific guidelines to evaluate different management schemes. For example, (Greenhill et al., 2003) determined typical ranges of the metrics in environmentally sustainable localities for an extensive suburban area on the southwest edge of London using multispectral IKONOS-2 imagery. The spatial distributions of the metrics provide new insight into landscape structure, which can be exploited in land use planning and in the construction of empirical spatial planning heuristics for sustainable urban development. If researchers determine the critical values or ranges of the landscape metrics where landscape retains its identity or there is positive effect on biodiversity then landscape metrics can be extremely useful indicators for measuring the effectiveness of the management schemes.
 |
 |
 |
An Overview of Their Use in Landscape Research",
Living Rev. Landscape Res., 3 (2009), 1, doi:10.12942/lrlr-2009-1, URL (accessed <date>): http://lrlr.landscapeonline.de/lrlr-2009-1.
 This work is licensed under a Creative Commons License.
© The author(s), except where otherwise noted.
This work is licensed under a Creative Commons License.
© The author(s), except where otherwise noted.